Окисление жирных кислот в организме. β-Окисление в митохондриях, окисление жирных кислот с нечетным числом углеродных атомов
НАВИГАЦИЯ ПО СТРАНИЦЕ
легко понять и запомнить
β-окисление жирных кислот в митохондриях
Если в ЖК с количеством атомов С до 10, она свободно попадает в матрикс митохондрии и там активируется в ацил-КоА.
Если ЖК с количеством атомов С больше 10, то активация происходит в цитозоле, и такой ацил-КоА диффузно (свободно, беспрепятственно) попадает через наружную мембрану в ММП, а сам через внутреннюю в матрикс попасть не может.
Но на наружной поверхности ВММ находится карнитин. Фермент карнитин-ацилтрансфераза I (КАТ I) связывает карнитин и ацил-КоА в ацил-карнитин, который легко проходит через ВММ:
ацил-КоА + карнитин ацил-карнитин + КоА-SH.
А уже в матриксе, на внутренней поверхности ВММ, КАТ II отсоединяет от ацил-карнитина, возвращает ему КоА:
КоА-SH + ацил-карнитин карнитин + ацил-КоА.
Реакции:

β-окисление жирных кислот
Комментарии:
ДГ отдает ФАДу протоны водорода, и между α- и β-атомами образуется С=С связь. Затем гидратаза образовывает ОН-группу, которую ДГ окисляет, отдав водороды НАДу. После чего тиолаза отрезает ацетил-КоА, а к концу присоединяется новый КоА и процесс идет до С4, после которого образуется 2 ацетил-КоА, и оба уйдут в цикл Кребса.
За 1 виток выходит:
1 восстановленный ФАД∙Н2 (уходит в дыхательную цепь – 1,5 АТФ);
1 восстановленный НАДН∙Н+ (уходит в дыхательную цепь – 2,5 АТФ);
1 ацетил-КоА (уходит в цикл Кребса – 10 АТФ).
Вывод: за один виток ЖК укорачивается на 2 атома С.
Расчет энергетического баланса β-окисления
Если количество С четное (Миристиновая к-та)
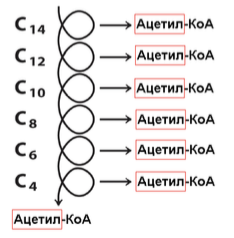
Получаем: 6 витков; 7 ацетил-КоА (по 10 АТФ); 6 ФАД∙ (по 1,5 АТФ); 6 НАДН∙Н+ (по 2,5 АТФ).
Итого: 94 – 2 (в реакции активации мы теряем 2 АТФ) = 92 АТФ.
Если количество С нечетное (Пентадекановая к-та)
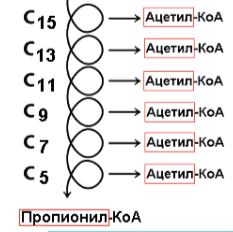
Получаем: 6 витков; 6 ацетил-КоА (по 10 АТФ); 1 пропионил-КоА (25 АТФ); 6 ФАД∙ (по 1,5 АТФ); 6 НАДН∙Н+ (по 2,5 АТФ).
Итого: 109 – 3 (в реакции активации мы теряем 2 АТФ + 1 АТФ описан ниже) = 106 АТФ.
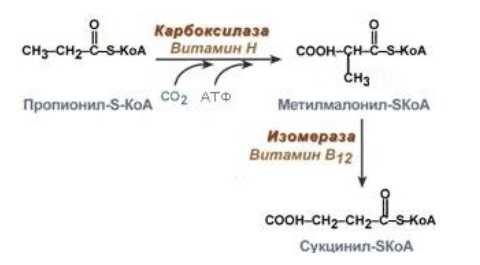
В этом случае кислота обычным способом окисляется до С3, остаеся пропионил-КоА. Он богат энергией, но в таком виде в цикл Кребса ему не попасть (нет такого субстрата), поэтому к нему присоединяется еще один углерод – карбоксилирование (тратится еще одна АТФ), после чего сможем получить сукцинил-КоА. Присоединить атом С получается только в середину цепи, поэтому нужна еще изомераза:
β-окисление ненасыщенных ЖК
Проходит до места двойной связи. Есть 2 выхода:
С=С связь оказалась в положении 3,4-цис, т.е. отходит от γ-атома. Изомераза перенесет С=С связь в положение 2,3-транс. Теперь она отходит от β-атома, что и требуется в процессе.
Можно просто насытить двойную связь с помощью редуктазы, потратив на это один из НАДФН∙Н+, после чего окисление продолжается дальше.
В первом случае мы недополучим ФАД∙Н2 (т.е. минус 1,5 АТФ), а во втором – используем один из полученных восстановленных НАДН∙Н+ (т.е. минус 2,5 АТФ). Это нужно учесть при расчете энергетического баланса.
Регуляция:
стимул – интенсивная мышечная работа (физическая нагрузка);
тормозится при увеличении соотношений (принцип обратной связи): ↑ ацетил-КоА/ацил-КоА; ↑ НАДН∙Н+/НАД+; ↑ ФАД∙Н2/ФАД+;
малонил-КоА – ингибитор КАТ I;
избыток ацетил-КоА ингибирует ПВДГК → ↓ гликолиз;
↑ гликолиз → накопление малонил-КоА → ингибир. КАТ I → ↓ β-окисление.
Недостаток ацил-КоА-ДГ – 10% внезапной младенческой смерти:
не идет β-окисление → основным топливом становится глюкоза, запасы гликогена у ребенка малы → гипогликемия → кома.
Вопрос: сколько АТФ мы получим за счет окисления трипальмитоилглицерола (ТАГ)?
Решение: разберем ТАГ на составляющие: глицерол + 3 пальмитата.
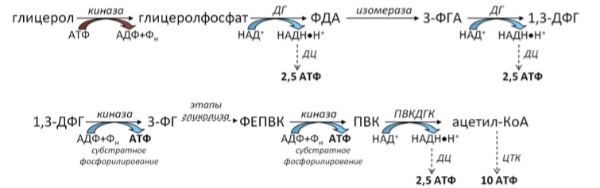
Всего: 2,5*3 + 2 + 10*1 = 18,5 АТФ
Пальмитиновая кислота (С15Н31СООН, С16):
Получаем 7 витков; 8 ацетил-КоА (по 10 АТФ); 7 ФАД∙ (по 1,5 АТФ); 7 НАДН∙Н+ (по 2,5 АТФ)
Всего: 80 + 10,5 + 17,5 = 106 АТФ
Ответ: 318 + 18,5 = 336,5 АТФ.
В пероксисомах окисляются:
ЖК с количеством С больше 18;
с разветвленным радикалом;
дикарбоновые.
Особенности β-окисления в пероксисоме:
в первой реакции ДГ образуется не ФАД∙, а , которую потом разрушит каталаза;
до С8, потом – в митохондрию, т.к. ацил-КоА-ДГ в пероксисоме последний раз может подействовать максимум на С10.
