Происхождение и эволюция человека
НАВИГАЦИЯ ПО СТРАНИЦЕ
легко понять и запомнить

Доказательства эволюции
Палеонтологические доказательства
Как правило, остатки растений и животных разлагаются и исчезают без следа. Но иногда биологические ткани замещаются минеральными веществами, и образуются окаменелости. Обычно находят окаменевшие кости или раковины, то есть твердые части живых организмов.
Иногда находят отпечатки следов животных или следы их жизнедеятельности. Еще реже находят животное целиком — вмороженным в лед в районах современной вечной мерзлоты, попавшим в окаменевшую позже смолу древних растений (янтарь) или в другую естественную смолу — асфальт.
Изучением ископаемых остатков занимается палеонтология.
Как правило, осадочные породы залегают слоями, поэтому более глубокие слои содержат окаменелости из более раннего периода. А значит, сравнивая ископаемые формы из последовательных напластований, можно делать выводы об основных направлениях эволюции живых организмов. Для оценки возраста окаменелостей используются многочисленные методы геохронологии.
При взгляде на палеонтологическую летопись можно сделать вывод, что жизнь на Земле существенно менялась. Чем глубже в прошлое мы смотрим, тем меньше видим общего с современной биосферой.
Первые прокариоты появляются приблизительно 3,5 млрд лет назад. Первые одноклеточные эукариоты появляются 2,7—1,75 млрд лет назад.
Примерно через миллиард лет, 840 млн лет назад, в палеонтологической летописи появляются первые многоклеточные животные. В период «кембрийского взрыва», 540—530 млн лет назад, за геологически короткий промежуток времени, в геологической летописи появляются остатки представителей большинства современных типов, имеющих скелеты, а еще через 15 млн лет — первые примитивные позвоночные, похожие на современных миног.
Челюстноротые рыбы появляются 410 млн лет назад, насекомые — 400 млн лет назад, и еще 100 млн лет на суше господствуют папоротниковидные, а основными группами наземной фауны остаются насекомые и земноводные.
С 250 по 65 млн лет назад на Земле господствующее положение «верховных хищников» и крупных травоядных занимали динозавры и другие рептилии, самыми распространенными растениями были саговники и другие группы голосеменных.
Первые ископаемые остатки цветковых появляются 140—130 млн лет назад, а начало их широкого распространения относится к середине мелового периода (около 100 млн лет назад). Наблюдаемая картина соответствует происхождению всех видов от одноклеточных организмов и не имеет других научных объяснений.
Известное доказательство эволюции — наличие так называемых промежуточных форм, то есть организмов, сочетающих в себе характерные признаки разных видов.
Как правило, говоря о промежуточных (или «переходных») формах имеют в виду ископаемые виды, хотя промежуточные виды не всегда вымирают. На основе филогенетического древа теория эволюции предсказывает, какие промежуточные формы могут быть найдены, а какие — нет.
В соответствии с научным методом, сбывшиеся предсказания подтверждают теорию. Например, зная строение скелета динозавров и произошедших из них птиц, можно предсказать некоторые особенности переходной формы между ними. Можно прогнозировать возможность найти останки животных, имеющих рептильный план строения, но с перьями, или остатки животных, подобных птицам, но с зубами или с длинными хвостами со скелетом из несросшихся позвонков. При этом можно предсказать, что не будут найдены переходные формы между птицами и млекопитающими, например, ископаемые млекопитающие с перьями или подобные птицам ископаемые с костями среднего уха как у млекопитающих.
Вскоре после публикации «Происхождения видов» были обнаружены остатки археоптерикса — промежуточной формы между рептилиями и птицами.
Для археоптерикса характерно дифференцированное оперение (типичная птичья черта), а по строению скелета он слабо отличался от динозавров из группы компсогнатов. У него были когти на передних конечностях, зубы и длинный хвост со скелетом из несросшихся позвонков, а предполагаемые уникальные «птичьи» особенности скелета впоследствии были выявлены у ряда других нептичьих динозавров. Позднее были найдены и другие переходные формы между продвинутыми рептилиями и птицами.

Известно множество других переходных форм, в том числе от беспозвоночных к рыбам, от рыб к четвероногим, от земноводных к рептилиям и от рептилий к млекопитающим.
В тех случаях, когда палеонтологическая летопись оказывается особенно полна, появляется возможность построить так называемые филогенетические ряды, то есть ряды видов (родов и т. д.), последовательно сменяющих друг друга в процессе эволюции.

Сравнительно-анатомические доказательства
В ходе эволюции каждый новый организм не проектируется с нуля, а получается из старого за счет последовательности небольших изменений. У образовавшихся таким образом структур есть ряд характерных особенностей, указывающих на их эволюционное происхождение. Сравнительно-анатомические исследования позволяют выявить такие особенности.
В частности, эволюционное происхождение исключает возможность целенаправленного заимствования удачных конструкций у других организмов. Поэтому у различных не близкородственных видов для решения схожих задач используются различные органы.
Например: крыло бабочки и крыло птицы развиваются из разных зародышевых листков, крылья птиц представляют собой видоизмененные передние конечности, а крылья бабочки — складки хитинового покрова.
Противоположная ситуация наблюдается у близкородственных видов: для совершенно разных задач используются схожие по строению органы.
Например: передние конечности позвоночных выполняют самые разные функции, но при этом имеют общий план строения, занимают сходное положение и развиваются из одних и тех же зачатков.
Сходство строения крыла летучей мыши и лапы крота невозможно объяснить с точки зрения полезности. В то же время, теория эволюции дает объяснение: единую структуру конечности четвероногие позвоночные унаследовали от общего предка.
Гомологичные органы
Передние конечности млекопитающих представляют собой пример гомологичных органов, так как происходят из одних и тех же тканей зародыша по сходному механизму, состоят из одного и того же набора костей, но при этом выполняют самые разные функции.
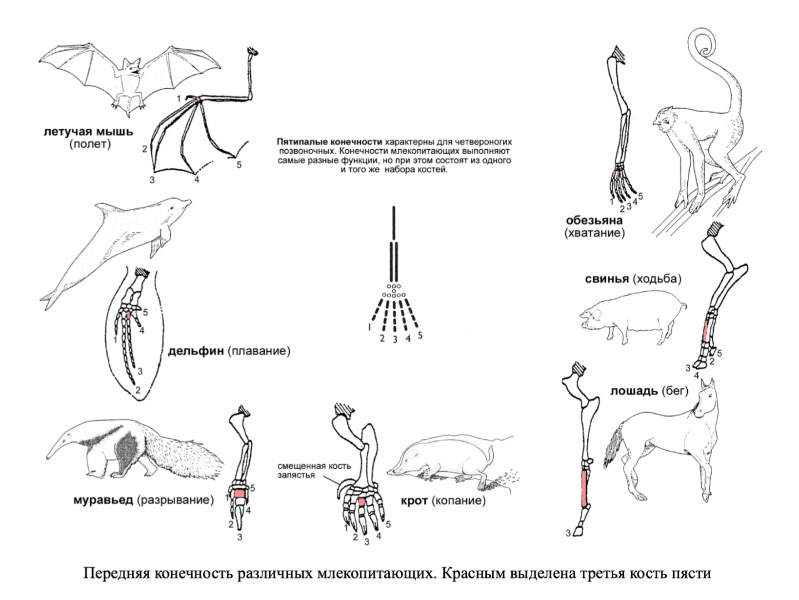
Более того, прослеживается гомология пятипалой конечности и плавников некоторых ископаемых видов кистеперых рыб, от которых, по всей видимости, произошли первые земноводные. Конечности четвероногих различаются по форме и приспособлены к выполнению самых различных функций в самых разных условиях.
При этом все эти конечности содержат сходный набор костей с одним и тем же относительным расположением. Единство структуры не может быть объяснено с точки зрения полезности, так как конечности используются для совершенно разных целей.
Аналогичные органы
Внешне схожие органы или их части, происходящие из различных исходных зачатков и имеющие неодинаковое внутреннее строение, называются аналогичными. Внешнее сходство возникает в ходе конвергентной эволюции, то есть в ходе независимого приспособления к сходным условиям существования.
Примеры:
Крылья птиц — видоизмененные передние конечности, крылья насекомых — складки хитинового покрова.
Жабры рыб — образования, связанные с внутренним скелетом, жабры многих ракообразных — выросты конечностей.
Обтекаемая форма тела у водных млекопитающих — китов, дельфинов — и у рыб.
Колючки барбариса, кактуса — видоизмененные листья, колючки боярышника развиваются из побегов.
Усики винограда (образующиеся из побегов) и усики гороха (видоизмененные листья).
Полное отсутствие целенаправленного заимствования удачных конструкций отличает эволюцию от сознательного проектирования.
Например: перо — это удачная конструкция, помогающая при полете, но у млекопитающих (в том числе и у летучих мышей) перья отсутствуют; жабры чрезвычайно полезны для водных животных, но у млекопитающих (таких как китообразные) они отсутствуют.
Чтобы опровергнуть теорию эволюции, достаточно обнаружить перья или жабры у какого-либо вида млекопитающих.
Рудименты
Рудиментами называются органы, утратившие свое основное значение в процессе эволюционного развития организма.
Рудименты также можно определить независимо от эволюционного учения как структуры, редуцированные и обладающие меньшими возможностями по сравнению с соответствующими структурами в других организмах. Если рудимент и оказывается функциональным, то он выполняет относительно простые или малозначимые функции с помощью структур, очевидно предназначенных для более сложных целей. Хотя многие рудиментарные органы совершенно не функциональны, отсутствие функций не является необходимым условием для рудиментарности. Такие органы крайне распространены в природе.
Примеры рудиментов:
Глаза у некоторых пещерных и роющих животных, таких как протей, слепыш, крот, астианакс мексиканский (слепая пещерная рыба). Часто глаза скрыты под кожей.
Малая берцовая кость у птиц.
Остатки волосяного покрова и тазовых костей у некоторых китообразных.
У некоторых змей, в том числе у питона, имеются кости задних конечностей. Эти кости не крепятся к позвоночнику и относительно свободно перемещаются в брюшной полости.
У многих видов жуков крылья лежат под сросшимися надкрыльями.
У человека к рудиментам в частности относятся хвостовые позвонки, волосяной покров туловища, ушные мышцы, бугорок ушной раковины, зубы мудрости, монгольская складка (остатки третьего века).
Червеобразный отросток слепой кишки (аппендикс) у некоторых травоядных животных используется для переваривания растительной пищи и имеет большую длину. Например, у коалы длина аппендикса составляет от 1 до 2 метров. Аппендикс человека имеет длину от 2 до 20 сантиметров и не участвует в расщеплении пищи. Вопреки распространенному убеждению, наличие у аппендикса второстепенных функций не означает, что он не является рудиментом.

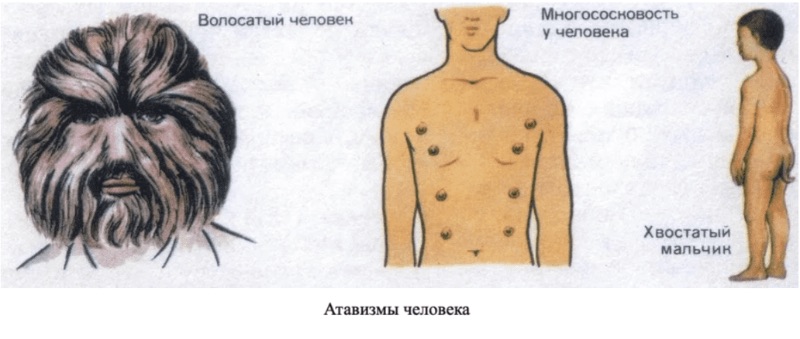
Атавизмы
Атавизмом называется появление у особи признаков, свойственных отдаленным предкам, но отсутствующих у ближайших. Появление атавизмов объясняется тем, что гены, отвечающие за данный признак, сохранились в ДНК, но в норме не формируют структуры, типичные для предков.
Примеры атавизмов:
Хвостовидный придаток у человека.
Сплошной волосяной покров на теле человека.
Добавочные пары молочных желез (многососковость).
Задние конечности у китообразных.
Задние ноги у змей.
Дополнительные пальцы у лошадей.
Возобновление полового размножения у ястребинки волосистой и у клещей семейства Crotoniidae.
Эмбриологические доказательства
У всех позвоночных животных наблюдается значительное сходство зародышей на ранних стадиях развития: у них похожая форма тела, есть зачатки жаберных дуг, имеется хвост, один круг кровообращения и т. д. (закон зародышевого сходства Бэра).
Однако по мере развития сходство между зародышами различных систематических групп постепенно уменьшается, и начинают преобладать черты, свойственные их классам, отрядам, семействам, родам и, наконец, видам.
Эволюционные изменения могут касаться всех фаз онтогенеза, то есть могут приводить к изменениям не только зрелых организмов, но и эмбрионов, даже на первых этапах развития. Тем не менее, более ранние фазы развития должны отличаться большим консерватизмом, чем более поздние, так как изменения на более ранних этапах развития, в свою очередь, должны привести к большим изменениям в процессе дальнейшего развития. Например, изменение типа дробления вызовет изменения в процессе гаструляции, равно как и во всех следующих стадиях. Поэтому изменения, проявляющиеся на ранних этапах, гораздо чаще оказываются летальными, чем изменения, касающиеся более поздних периодов онтогенеза.
Таким образом, ранние стадии развития изменяются относительно редко, а значит, изучая эмбрионы разных видов, можно делать выводы о степени эволюционного родства.
Есть многие другие примеры того, как эволюционная история организма проявляется в ходе его развития.
У эмбрионов млекопитающих на ранних стадиях есть жаберные мешки, неотличимые по строению от жаберных мешков у водных позвоночных. Это объясняется тем, что предки млекопитающих жили в воде и дышали жабрами. Разумеется, жаберные мешки зародышей млекопитающих в ходе развития превращаются не в жабры, а в структуры, возникшие в ходе эволюции из жаберных щелей или стенок жаберных карманов, такие как евстахиевы трубы, среднее ухо, миндалины, паращитовидные железы и тимус.
У эмбрионов многих видов змей и безногих ящериц (например, веретеницы ломкой) развиваются зачатки конечностей, но затем они рассасываются. Аналогично у китов, дельфинов и морских свиней нет задних конечностей, но у эмбрионов китообразных начинают расти задние ноги, развиваются кости, нервы, сосуды, а затем все эти ткани рассасываются.
Биогенетический закон Бэра-Геккеля-Мюллера: процесс эмбрионального развития (онтогенез) зародыша данного вида есть краткое повторение исторического развития данного вида (филогенез). Не стоит относиться к нему как к научной догме, т. к. существует большое количество фактов, ему не соответствующих.

Молекулярно-генетические доказательства
Живые организмы имеют удивительно сходный химический состав.
У всех форм жизни, за исключением некоторых вирусов, хранителем наследственной информации является ДНК. Кодируемые ею белки играют первостепенную роль в осуществлении различных процессов жизнедеятельности.
Поставщиком энергии для протекания этих процессов служит, главным образом, АТФ.
Для синтеза биомолекул живые организмы используют преимущественно одни и те же структурные компоненты. Например, ДНК построена из четырех основных типов нуклеотидов, а белки — из 20 видов аминокислот.
Высокая степень сходства проявляется не только в строении биологических молекул, но и в их функционировании.
Так, у живых организмов сходным образом протекают процессы энергетического обмена и реакции матричного синтеза. Реализация наследственной информации, зашифрованной с помощью универсального генетического кода, у всех организмов осуществляется благодаря транскрипции и трансляции, с участием мРНК, тРНК и рРНК. Эти и многие другие факты доказывают родство живых организмов и единство происхождения жизни на Земле.
Биохимические доказательства
Носителем наследственной информации во всех клетках являются молекулы ДНК, у всех известных организмов в основе размножения — репликация этой молекулы.
В ДНК всех организмов используются 4 нуклеотида (аденин, гуанин, тимин, цитозин), хотя в природе встречаются не менее 102 различных нуклеотидов. Кроме того, в природе встречается 390 различных аминокислот, но белки всех организмов составляются из одного и того же набора, в котором всего 22 аминокислоты. При этом возможно 1,4⋅1070 различных информационно эквивалентных генетических кодов, использующих те же самые кодоны и аминокислоты.
Если не учитывать эволюционное происхождение всех организмов от общего предка, то ничто не мешает каждому виду иметь собственный генетический код. Такое положение вещей было бы крайне выгодным, так как при этом исключалось бы преодоление вирусами межвидовых барьеров. Тем не менее, ничего подобного не наблюдается, и теория эволюции исключает такую возможность: изменения генетического кода приводят к изменению большинства белков организма, такая мутация практически всегда оказывается летальной, поэтому код не мог значительно измениться со времен последнего общего предка, что гарантирует его универсальность.
Также можно отметить крайне похожие пути метаболизма в клетках всех организмов.
Так, гликолиз у всех эукариот и у большинства прокариот проходит за 10 одинаковых шагов, в одной и той же последовательности, с использованием одних и тех же десяти ферментов (при том что возможны тысячи различных, но термодинамически эквивалентных путей гликолиза). У всех изученных видов основным переносчиком энергии в клетке является аденозинтрифосфат (АТФ), хотя эту роль могли бы выполнять сотни других молекул.
Биогеографические доказательства
Биогеография — наука, изучающая закономерности географического распространения животных и растений, а также характер фауны и флоры отдельных территорий.
Если два вида сравнительно недавно произошли от одной популяции, то они, как правило, обитают недалеко от ареала этой исходной популяции, а значит и недалеко друг от друга.
Таким образом, географическое распределение видов должно быть совместимо с филогенетическим деревом. Если не принимать во внимание теорию эволюции, то разумно предположить, что виды живут в наиболее подходящих для них условиях. Теория эволюции же предсказывает, что должно быть много благоприятных для вида мест, в которых представители вида тем не менее отсутствуют, в связи с наличием географических барьеров.
Именно так дело и обстоит на практике. Среди млекопитающих Австралии преобладают сумчатые. Плацентарные млекопитающие представлены китообразными, ластоногими и рукокрылыми (которые могли перебраться в Австралию сравнительно легко), а также грызунами, которые появляются в палеонтологической летописи в миоцене, когда Австралия приблизилась к Новой Гвинее.
При этом природные условия Австралии благоприятны и для других видов млекопитающих. Например, завезенные на континент кролики быстро размножились, широко расселились и продолжают вытеснять аборигенные виды.
В Австралии и Новой Гвинее, на юге Южной Америки и в Африке встречаются нелетающие бескилевые птицы, свистуны (зубастые жабы) и двоякодышащие, в других частях света они отсутствуют. Условия обитания в пустынях Африки, Америки и Австралии очень похожи, и растения из одной пустыни хорошо растут в другой. Тем не менее, кактусы были обнаружены только в Америке.
Многие африканские и австралийские суккуленты (то есть растения, имеющие специальные ткани для запасания воды) внешне напоминают кактусы за счет конвергентной эволюции, но относятся к другим порядкам.
Морские обитатели восточных и западных берегов Южной Америки различны, за исключением некоторых моллюсков, ракообразных и иглокожих, но на противоположных берегах Панамского перешейка живет около 30% одних и тех же видов рыб, что объясняется недавним возникновением перешейка (около 3 млн лет назад).
На большинстве океанических островов (то есть островов, которые никогда не были соединены с материком) отсутствуют наземные млекопитающие, земноводные и другие животные, не способные преодолевать значительные водные преграды. Видовой состав фауны таких островов беден и является результатом случайного заноса некоторых видов, обычно птиц, ящериц и насекомых.
Географическое распределение видов в прошлом, которое можно частично восстановить по ископаемым остаткам, также должно соответствовать филогенетическому дереву.
Теория дрейфа материков и теория эволюции позволяют предсказать, где следует искать те или иные ископаемые остатки.
Первые окаменелости сумчатых найдены в Северной Америке, их возраст составляет около 80 млн лет. 40 млн лет назад сумчатые уже были распространены в Южной Америке, но в Австралии, где они сейчас доминируют, сумчатые появились только около 30 млн лет назад. Теория эволюции предсказывает, что австралийские сумчатые произошли от американских.
Согласно теории дрейфа материков, 30—40 млн лет назад Южная Америка и Австралия еще оставались частью Гондваны, крупного континента в Южном полушарии, а между ними находилась будущая Антарктида. На основании двух теорий исследователи предсказали, что сумчатые мигрировали из Южной Америки в Австралию через Антарктиду 30—40 млн лет назад. Это предсказание сбылось: начиная с 1982 года на острове Сеймур, расположенном недалеко от Антарктиды, были найдены более десяти ископаемых сумчатых возрастом 35—40 млн лет.
Наиболее близкие родственники современных людей — гориллы и шимпанзе — обитают в Африке. Исходя из этого, в 1872 году Чарльз Дарвин предположил, что и древних предков человека следует искать в Африке.
Многие исследователи последовали совету Дарвина, и начиная с 1920-х годов в Африке было найдено множество промежуточных форм между человеком и человекообразными обезьянами. Если бы ископаемых австралопитеков обнаружили, например, в Австралии, а не в Африке, то представления об эволюции гоминид пришлось бы пересматривать.
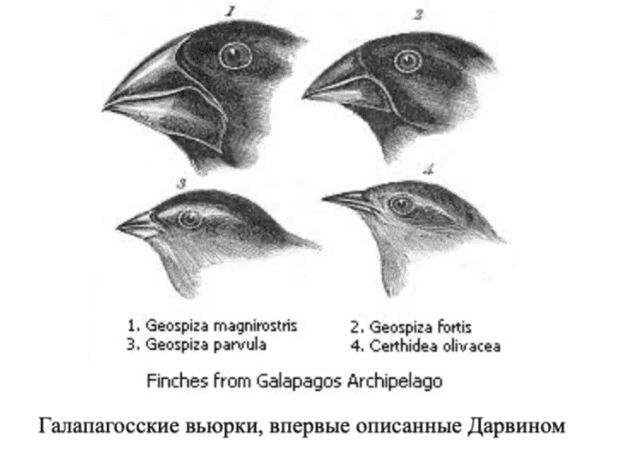
Во время кругосветного путешествия на корабле «Бигль» Чарльз Дарвин описал 13 видов вьюрков, обитающих на Галапагосских островах.
Наблюдение за этими птицами натолкнуло Дарвина на идею происхождения видов за счет изменчивости и естественного отбора.
Все галапагосские вьюрки происходят от общего предка, случайно попавшего сюда из Южной Америки. От предковой формы, питавшейся семенами, произошли три группы птиц: семяноядные земляные вьюрки, насекомоядные древесные вьюрки и славковые вьюрки, которые тоже питаются мелкими насекомыми. В результате приспособления к разным источникам пищи, вьюрки стали сильно отличаться друг от друга строением клюва. Три обычных вида земляных вьюрков — большой, средний и малый — встречаются на большинстве островов. В этом случае они хорошо различаются по размерам клюва и, соответственно, по величине предпочитаемых семян. На одном из островов обитает лишь средний земляной вьюрок, и здесь у птиц клювы меньше — в отсутствие конкурента средний земляной вьюрок занимает отчасти нишу малого вьюрка.
Реликты различают по геологическому возрасту флор и фаун прошлых эпох, от которых они происходят.
Филогенетические реликты (живые ископаемые) — ныне существующие виды растений и животных, которые относятся к крупным таксонам, почти полностью вымершим десятки или сотни миллионов лет назад.
Примеры: латимерия, гаттерии, крокодилы, однопроходные млекопитающие, некоторые сумчатые (например, опоссумы), споровые растения, гинкго, саговник, цианобактерии и др.
Географические реликты — организмы, сохранившиеся в данном регионе как остаток флор и фаун минувших геологических эпох, когда условия существования в данном регионе заметно отличались от современных.
Эндемики — специфическая составная часть какой-либо флоры, фауны.
К эндемикам относятся виды, роды, семейства или другие таксоны животных и растений, представители которых обитают на относительно ограниченном ареале, представлены небольшой географической областью. Пример: байкальская нерпа.
