Лигандзависимые и другие виды ионных каналов. Их строение, физиологическая роль и механизмы внутриклеточной передачи сигнала
НАВИГАЦИЯ ПО СТРАНИЦЕ
легко понять и запомнить
Ионный канал — несколько субъединиц (интегральных мембранных белков, содержащих трансмембранные сегменты, каждый из которых имеет а-спиральную конфигурацию), обеспечивающих перенос ионов через мембрану.
Каналообразующая молекула белка располагается в цитоплазматической мембране так, что ее трехмерная пространственная структура формирует устья канала, обращенные к наружной и внутренней сторонам мембраны, пору, заполненную водой, и «ворота». Последние образуются участком пептидной цепи, который может легко изменять свою конформацию и определять открытое или закрытое состояние канала. От размеров поры и ее заряда зависят селективность и проницаемость ионного канала. Проницаемость канала для данного иона определяется также его размерами, величиной заряда и гидратной оболочкой.
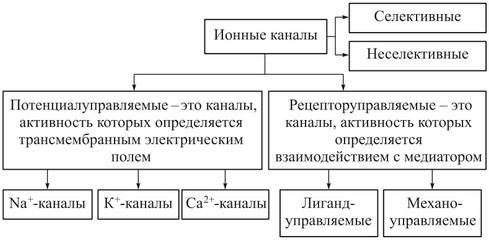
Типы ионных каналов
пассивные ионные каналы, или каналы покоя;
каналы щелевых контактов;
каналы, состояние которых (открытое или закрытое) контролируется влиянием на их воротный механизм механических факторов (механочувствительные каналы), разности потенциалов на мембране (потенциалзависимые каналы) или лигандов, связывающихся с каналообразующим белком на внешней или внутренней стороне мембраны (лигандзависимые каналы).
Пассивные каналы
Могут быть открытыми (активными) в покоящихся клетках, т.е. в отсутствие каких-либо воздействий (поэтому пассивные).
Не являются строго селективными, и через них мембрана клетки может «протекать» для нескольких ионов, например К+ и CI-, К+ и Na+. Поэтому иногда эти каналы называют каналами утечки.
Играют важную роль в возникновении и поддержании на цитоплазматической мембране клетки мембранного потенциала покоя.
Представлены в цитоплазматических мембранах нервных волокон и их окончаний, клеток поперечно-полосатой, гладкой мускулатуры, миокарда и других тканей.
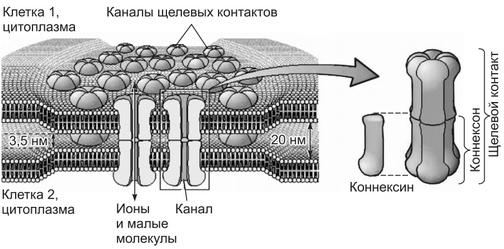
Каналы щелевых (плотных) контактов клеток
Каналы щелевых контактов формируются в области контакта двух соседних клеток, очень близко прилежащих друг к другу. В мембране каждой контактирующей клетки шесть белковых субъединиц, получивших название коннексины, формируют гексагональную структуру, в центре которой образуется пора или ионный канал — коннексон.
Зеркальная структура образуется в месте контакта в мембране соседней клетки, и ионный канал между ними становится общим. Через такие ионные каналы из клетки в клетку могут перемещаться различные минеральные ионы, в том числе ионы Са2+, а также низкомолекулярные органические вещества. Каналы щелевых контактов клеток обеспечивают передачу информации между клетками миокарда, гладких мышц, сетчатки глаза, нервной системы.
Механочувствительные каналы
Состояние проницаемости этих каналов изменяется при механических воздействиях на мембрану, вызывающих нарушение структурной упаковки молекул в мембране и ее растяжение. Эти каналы широко представлены в ЦПМ механорецепторов сосудов, внутренних органов, кожи, поперечно-полосатых мышц, гладких миоцитов.
Потенциалзависимые каналы

Их состояние управляется силами электрического поля, создаваемого величиной разности потенциалов на мембране.
Потенциалзависимые ионные каналы обычно являются высокоселективными и играют решающую роль в возникновении возбуждения (генерации потенциалов действия), передаче информации по нервным волокнам в виде электрических сигналов, инициировании и регуляции сокращения мышц.
Могут находиться в неактивном (закрытом), активном (открытом) и инактивированном состояниях, которые контролируются положением активационных и инактивационных ворот, зависящим от разности потенциалов на мембране.
В покоящейся клетке потенциалзависимый канал находится обычно в закрытом состоянии, из которого он может быть открыт или активирован. Вероятность его самостоятельного открытия невысокая, и в состоянии покоя лишь небольшое количество этих каналов в мембране открыто. Уменьшение трансмембранной разности потенциалов (деполяризация мембраны) вызывает активацию канала, повышая вероятность его открытия. Функцию активационных ворот выполняет электрически заряженная аминокислотная группа, закрывающая вход в устье канала. Эти аминокислоты являются сенсором разности потенциалов на мембране; при достижении определенного (критического) уровня деполяризации мембраны заряженная часть молекулы сенсора смещается в сторону липидного микроокружения каналообразующей молекулы и ворота открывают вход в устье канала.
Канал становится открытым (активным) для перемещения через него ионов. Скорость открытия активационных ворот может быть низкой и очень высокой. По этому показателю потенциалзависимые ионные каналы делят на:
быстрые (например, быстрые потенциалзависимые натриевые каналы);
медленные (например, медленные потенциалзависимые кальциевые каналы).
Быстрые каналы открываются мгновенно (мкс) и остаются открытыми в среднем в течение 1 мс. Их активация сопровождается быстрым лавинообразным возрастанием проницаемости канала для определенных ионов.
Способностью изменять свою конформацию обладает другая часть пептидной цепи, представляющая собой аминокислотную последовательность в виде плотного шарика (клубка) на нити, располагающегося у выхода другого устья канала. При изменении знака заряда на мембране шарик закрывает выход из устья, канал становится непроницаемым (инактивированным) для иона. Инактивация потенциалзависимых ионных каналов может осуществляться с помощью других механизмов. Инактивация сопровождается прекращением перемещения ионов через канал и может протекать так же быстро, как активация, или медленно — в течение секунд и даже минут.
Реактивация канала – переход из состояния инактивации в исходное (закрытое) состояние. При этом происходит возвращение исходной пространственной конформации каналообразующего белка и положение ворот. Реактивация происходит при восстановлении разности потенциалов мембраны (реполяризации) до уровня, характерного для состояния покоя клетки. Будучи реактивированным, ионный канал вновь возвращается в состояние готовности к его повторному открытию. Реактивация потенциалзависимых каналов мембраны также может быть быстрой и медленной.
Представлены в мембранах афферентных, эфферентных нервных волокон, в мембранах поперечно-полосатых и гладких миоцитов.
Существуют вещества, способные блокировать работу этих каналов. Одним из первых среди них было описано вещество тетродотоксин — сильнейший яд, образующийся в организме рыбы фугу. Под его действием наблюдалась блокада потенциалзависимых натриевых каналов, а при его введении в организм животных отмечались потеря чувствительности, расслабление мышц, обездвиженность, остановка дыхания и смерть. Такие вещества получили название блокаторов ионных каналов. Среди них лидокаин, новокаин, прокаин — вещества, при введении которых в организм в небольших дозах развивается блокада потенциапзависимых натриевых каналов нервных волокон и блокируется передача в ЦНС сигналов от болевых рецепторов. Эти вещества широко используются в медицинской практике в качестве местных обезболивающих средств.
Перемещение ионов через ионные каналы не только является основой перераспределения зарядов на мембранах и формирования электрических потенциалов, но и может оказывать влияние на протекание многих внутриклеточных процессов. Такое влияние на экспрессию генов, контролирующих синтез каналообразующих белков, не ограничено только клетками возбудимых тканей, но имеет место во всех клетках организма.
Лигандзависимые ионные каналы
Они обычно формируются белковыми макромолекулами, которые могут одновременно выполнять функцию ионных каналов и рецепторную функцию по отношению к определенным лигандам. Поскольку одна и та же макромолекула может одновременно выполнять эти две функции, то за ними закрепились разные названия — например, синаптический рецептор или лигандзависимый канал.
Лигандзависимые ионные каналы открываются (активируются) при взаимодействии пептидной (рецепторной) цепи белковой молекулы с лигандом — веществом, к которому рецептор имеет высокое сродство.
: а неактивный; 6 — активированный")
Лигандзависимый ионный канал (никотинчувствительный ацетилхолиновый рецептор — н-ХР): а неактивный; 6 — активированный
Лигандзависимые ионные каналы обычно локализованы в постсинаптических мембранах нервных клеток и их отростков, а также мышечных волокон. Типичными примерами лигандзависимых ионных каналов являются каналы постсинаптических мембран, активируемые ацетилхолином, глутаматом, аспартатом, гамма-аминомасляной кислотой, глицином и другими синаптическими нейромедиаторами. Обычно название канала (рецептора) отражает тип нейромедиатора, который в естественных условиях является его лигандом. Так, если это каналы нервно-мышечного синапса, в котором используется нейромедиатор ацетилхолин, то используется термин «ацетилхолиновый рецептор», а если он является к тому же чувствительным к никотину, то его называют никотинчувствительным, или просто н-ацетилхолиновым, рецептором (н-холинорецептор).
Обычно постсинаптические рецепторы (каналы) избирательно связываются только с одним типом нейромедиатора. В зависимости от типа и свойств взаимодействующих рецептора и нейромедиатора каналы избирательно изменяют свою проницаемость для минеральных ионов, но при этом они не являются строго селективными каналами. Например, лигандзависимые каналы могут изменять проницаемость для катионов Na+ и К+ или для К+ и анионов СI+. Такая избирательность связывания лиганда и изменения ионной проницаемости генетически закреплена в пространственной структуре макромолекулы.
Если взаимодействие медиатора и рецепторной части макромолекулы, формирующей ионный канал, непосредственно сопровождается изменением проницаемости канала, то в течение нескольких миллисекунд это приводит к изменению проницаемости постсинаптической мембраны для минеральных ионов и величины постсинаптического потенциала. Такие каналы названы быстрыми и локализуются, например, в постсинаптической мембране аксодендритных возбуждающих синапсов и аксосоматических тормозных синапсов.
Существуют медленные лигандзависимые ионные каналы. В отличие от быстрых каналов их открытие опосредованно не прямым взаимодействием нейромедиатора с макромолекулой-рецептором, а цепочкой событий, включающих активацию G-белка, его взаимодействие с GTP, повышение уровня вторичных посредников во внутриклеточной передаче сигнала нейромедиатора, которые, фосфорилируя ионный канал, приводят к изменению его проницаемости для минеральных ионов и соответствующему изменению величины постсинаптического потенциала.
Особым типом являются каналы, локализованные в мембранах эндоплазматичсского ретикулума гладкомышечной клетки. Их лигандом является вторичный посредник внутриклеточной передачи сигнала ИТФ.
Лигандзависимые ионные каналы широко представлены в мембранных структурах, обеспечивающих синаптическую передачу сигналов от ряда сенсорных рецепторов в ЦНС; передачу сигналов в синапсах нервной системы; передачу сигналов нервной системы к эффекторным клеткам.
Уже отмечалось, что непосредственная передача команд нервной системы ко многим эффекторным органам осуществляется с помощью нейромедиаторов, активирующих лигандзависимые ионные каналы в постсинаптических мембранах. Однако их лигандами (агонистами или антагонистами) могут быть и вещества экзогенной природы, которые в ряде случаев используются в качестве лекарственных веществ.
Например, после введения в организм вещества диплацина, сходного по своей структуре с нейромедиатором ацетилхолином, последует продолжительное открытие лигандзависимых ионных каналов нервно-мышечных синапсов, которые перестают передавать нервные импульсы с нервных волокон к мышцам. Наступает релаксация скелетных мышц организма, которая может понадобиться во время проведения сложных хирургических операций. Диплацин и другие вещества, способные изменять состояние лигандзависимых ионных каналов и блокировать передачу сигналов в нервно-мышечных синапсах, получили название миорелаксантов.
Потенциалзависимые натриевые каналы
Они состоят из одной а-субъединицы, формирующей канал, и двух β-субъединиц, модулирующих ионную проницаемость и кинетику инактивации натриевых каналов.
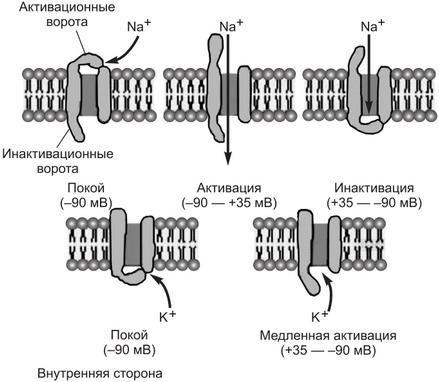
В потенциалзависимых натриевых каналах имеются два воротных механизма, один из них — активационный – обеспечивает открытие (активацию) канала при деполяризации мембраны, а второй — его инактивацию при перезарядке мембраны. Поскольку оба этих механизма быстро изменяют положение ворот канала, то потенциалзависимые натриевые каналы являются быстрыми ионными каналами и имеют определяющее значение для генерации потенциалов действия в возбудимых тканях и для их проведения по мембранам нервных и мышечных волокон.
Плотность распределения натриевых каналов в этих структурах различна. В миелинизированных нервных волокнах они сосредоточены главным образом в области перехватов Ранвье, где их плотность достигает около 10 000 каналов на квадратный микрон площади, а в немиелинизированных волокнах каналы распределены более равномерно с плотностью около 20 каналов на ту же площадь. Эти каналы практически отсутствуют в структуре мембран тела нервной клетки, в мембране нервных окончаний, непосредственно формирующих сенсорные рецепторы, и в постсинаптических мембранах эффекторных клеток.
Синтезированы и широко используются в медицине для блокады быстрых потенциалзависимых натриевых каналов вещества, получившие название анестетиков (новокаин, дикаин, лидокаин, совкаин, прокаин и др.). Анестезия при блокаде ими натриевых каналов достигается за счет устранения возможности генерации нервных импульсов в афферентных нервных волокнах и тем самым блокады проведения сигналов от сенсорных болевых рецепторов в ЦНС.
Обнаружено, что изменение структуры натриевых каналов может вести к развитию ряда заболеваний. Так, например, изменение структуры канала, контролируемого геном SCNlb, ведет к развитию генерализованных форм эпилепсии и судорог при повышении температуры тела (фебрильных судорог).
