Физиологические свойства сократительного миокарда. Потенциал действия клеток сократительного миокарда
НАВИГАЦИЯ ПО СТРАНИЦЕ
легко понять и запомнить
Отличия сердечной мышцы от скелетной:
Сокращение целостной мышцы подчиняется закону «все или ничего», т.е. при действии раздражителя пороговой силы отвечает максимальным сокращением. Дальнейшее увеличение силы не приводит к увеличению сокращения. Это свойство обусловлено наличием функционального синтиция (т.е. возбуждение может распространяться к другим клеткам миокарда).
Длительность сокращения больше, чем у одиночного сокращения скелетной мышцы (для Ж – 300-400 мс, для П – 100 мс). Длительность систолы зависит от ЧСС. При ее увеличении длительность систолы уменьшается.
Сердечная мышца находится в состоянии абсолютной рефрактерности в течение всей продолжительности возбуждения и не может сокращаться в ответ на новые воздействия, что предотвращает возникновение в ней тетанического сокращения.
Механизм запуска и самого сокращения кардиомиоцитов отличается тем, что поступление ионов Са в саркоплазму происходит не только из саркоплазматического ретикулума, но и из межклеточной жидкости, связанного с внутренней поверхностью, и из митохондрий.
На работу сердечной мышцы затрачивается АТФ, полученная в ходе окислительного фосфорилирования, что обеспечивает исключительную зависимость работы сердца и жизнеспособности миокарда от постоянной доставки кислорода и питаттельных веществ.
Сокращение происходит при поступлении ПД из ПСС, миокард не иннервируется волокнами соматической НС.
Величина поляризации покоя мембраны сократительных волокон миокарда равна -80-90 мВ (фаза 4), амплитуда их ПД (120 мВ) такие же, как и у скелетной, но длительность ПД примерно в 100 раз больше. Потенциал действия клеток рабочего миокарда состоит из:
фаза быстрой деполяризации (0 фаза);
фаза начальной быстрой реполяризации (1 фаза);
фаза медленной реполяризации (фаза плато, или 2 фаза) и фазы быстрой конечной реполяризации (3 фаза);
фаза покоя — (4 фаза).
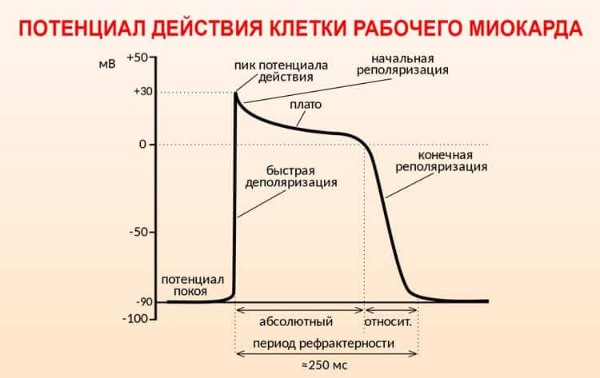
Фаза быстрой деполяризации создается активацией быстрых потенциалзависимых натриевых каналов, обеспечивающих резкое повышение проницаемости мембраны для ионов натрия, что приводит к возникновению быстрого входящего натриевого тока. Мембранный потенциал уменьшается от -90 мВ до +30 мВ, т.е. во время пика происходит изменение знака мембранного потенциала.
При достижении мембранного потенциала +30 мВ инактивируются быстрые натриевые каналы. Деполяризация мембраны вызывает активацию медленных натрий-кальциевых каналов. Поток ионов Са2+ внутрь клетки по этим каналам приводит к развитию плато ПД (фаза 2). В период плато клетка переходит в состояние абсолютной рефрактерности.
Затем происходит активация калиевых каналов. Выходящий из клетки поток ионов К обеспечивает быструю реполяризацию мембраны (фаза 3), во время которой медленные натрий-кальциевые каналы закрываются, что ускоряет процесс реполяризации.
Реполяризация мембраны вызывает постепенное закрывание калиевых и реактивацию натриевых каналов. В результате возбудимость миокардиальной клетки восстанавливается — это период так называемой относительной рефрактерности.
Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя (фаза 4).
